DNA folding and organization within a tiny nucleus has fascinated researchers for several decades. Recent studies have shown the importance of genome organization in the regulation of gene expression, DNA replication, recombination and cell division. Recent developments in ‘chromosome conformation capture’ (3C) technique have revealed details of genome organization particularly in the context of human diseases. Genomic rearrangements, a common feature of the cancer genome, often disorganized higher order of genome organization leading to inactivation of tumour suppressors or activation of oncogenes. However, unravelling the pathogenicity associated with 3D genome still remains a challenge, as internal structures and elements contributing for organization of the DNA within nucleus are yet to be understood. Importantly, how chromatin compartments/domains are spatially formed within the nucleus and to what extent changes in domains causes gene miss-expression, is still an open question. By using integrated genomics approaches, we wish to decode mechanisms responsible for chromatin folding in three-dimensional space and how these features are deregulated during cancer development.
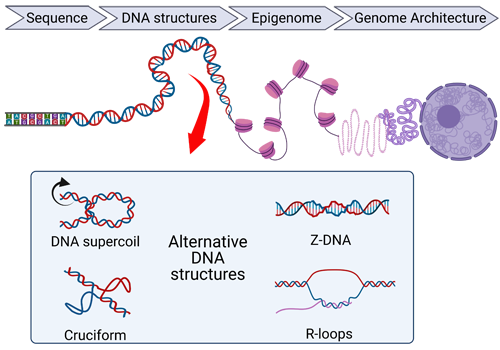
Our working hypothesis is that our genomic DNA is dynamic with regular deformation to its helical structure, generating supercoiling, bending and twisting. These mechanical and physical properties of chromatin along with alternative DNA structures (cruciform, Z-DNA, H-DNA), play a crucial role in dictating genome organization.
Based on our hypothesis, our research focuses on two major research areas:
DNA supercoil and chromatin organization
Objective here is to understand the role of super-helical behaviour of DNA in chromatin confirmation changes during cancer development.
DNA supercoil role in dictating RNA splicing and transcription control
Objective here is to explore chromatin influence on mRNA splicing, with particular emphasis on cancer associated alternative splicing.
Positions Available
The Genome Architecture lab at CDFD has openings for potential PhD students. Successful candidates will work on understanding DNA topology influence on (i) genome three-dimensional architecture (ii) splicing and transcription regulation. Our approach combines a great variety of genomics and molecular biology techniques with computational analysis using cancer cell lines and human primary tumours. We are looking for highly motivated candidates who wish to obtain hands-on skills in advanced genomics approaches including, Hi-C, SPRITE, ChIP-Seq, DRIP-Seq, RNA-seq and whole genome analysis.